36. 胎生期大脳におけるミクログリアの分布調節機構とその意義
研究の背景
私たちの脳を構成する細胞には、神経幹細胞を由来とするニューロン、アストロサイト、オリゴデンドロサイトといった神経系の細胞のほかにも、免疫系の細胞であるミクログリアが存在し、脳の機能構築を支えています。大人の脳におけるミクログリアの役割については、異物や死んだ細胞を貪食して除去することにより脳内の環境を整える1、神経回路が適切につくられるように監視する2などの役目のほか、神経変性疾患や感染症等の罹患時においても炎症応答や脳構造の修復に関わる3ことが知られます。一方で、胎生期での役割や存在意義はよくわかっていませんでした。
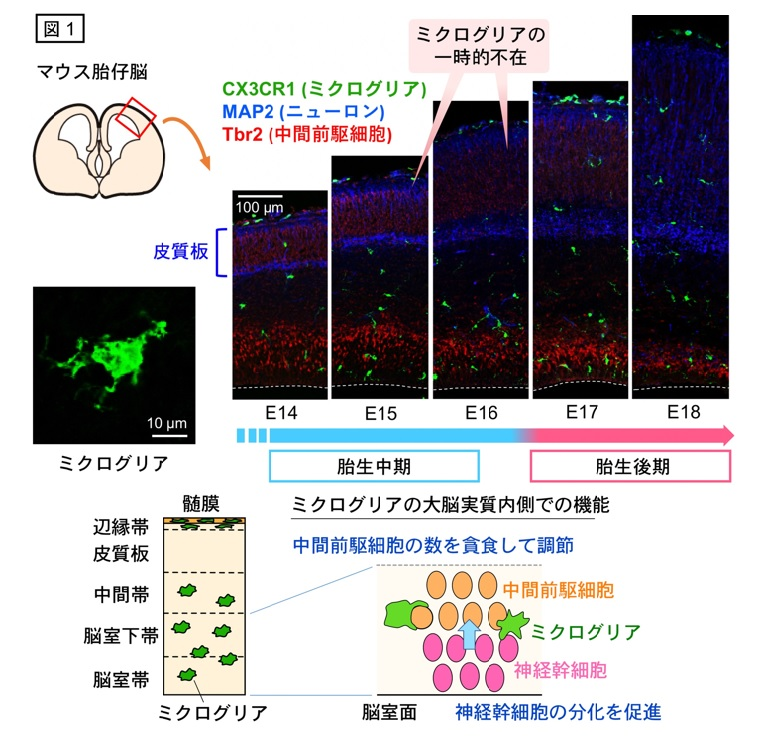
胎生期の大脳の発生は、神経系細胞の産生や分化に伴う移動が細やかに制御されて進んでいきます。一方で、ミクログリアも胎生早期から存在します。ミクログリアは脳を構成する全細胞のうち約1パーセント程度(胎生中期において)と少数ですが、細胞突起を遠くへ伸ばしながら脳実質内を広く動き回り、周辺の多数の神経系細胞と接することができます。私たちは、ミクログリアが神経幹細胞から中間前駆細胞への分化を促進し、脳実質内を広範囲に動くことによってその役目が効率的に果たされることを明らかにしました4。一方、海外グループの研究成果から、ミクログリアが貪食により神経系中間前駆細胞の数を調節していること5や、大脳基底核から移動し皮質板へと進入する介在ニューロンの配置に貢献していること6もわかってきています。
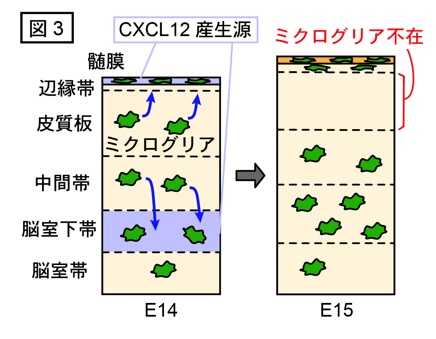
興味深いことに、ミクログリアは発生ステージの進行に伴い分布変化を示すことが知られています。すなわち、マウス胎生14日目(E14)までは大脳実質全体に均一に存在していますが、E15〜E16において皮質板と呼ばれる領域(分化を遂げたニューロンが積み重なる場所)から不在となります。そして、E17になると再び皮質板に分布するようになります(図1)。ミクログリアが本来存在する場所(脳室帯、脳室下帯、中間帯)における機能は上述のようなことが明らかとなってきている一方、胎生中期に皮質板から一時的に不在となるしくみや意義はこれまで問われていませんでした。そこで私たちは、マウス胎生期の大脳実質におけるミクログリアの分布変化のメカニズムと、脳発生過程におけるその生理学的意義について調べることにしました。
研究成果
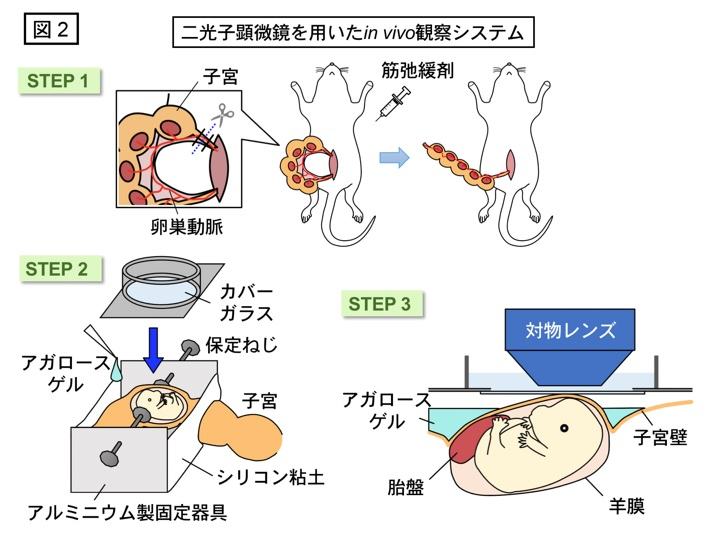
はじめに、ミクログリアがいかにしてE15までに皮質板から不在となるのかを調べるために、切り出した脳の組織片を培養しながら3次元的に細胞の動きを観察するスライス培養法、及び、二光子顕微鏡を用いてマウスが生きたままの状態で脳内の現象を観察する手法(図2)を用いて、ミクログリアの動態を経時的に見てみました。その結果、E14においてミクログリアが脳実質内を活発に移動する様子を捉えました。重要な点として、ミクログリアがもともと存在していた位置に依存して移動の方向性が決まり、皮質板内に存在するミクログリアは髄膜方向(外側)へと進み辺縁帯へと蓄積すること、また、中間帯に存在する細胞は脳室下帯の方向(内側)へ向かうという、両方向性に移動する性質が認められました。
次に、このミクログリアの両方向性の移動を制御する分子を探索しました。私たちは、ケモカインの一種であるCXCL12がE14の大脳において髄膜と脳室下帯で特異的に産生されることに着目しました。
ミクログリアはCXCL12の受容体であるCXCR4を発現することから、ミクログリアの移動を促すシステムとしてCXCL12-CXCR4を候補として考え、検証しました。 Cxcr4を欠損した遺伝子改変マウスを用いてミクログリアのライブ観察をおこなったところ、正常マウスで認められた両方向性の移動の特徴が認められなくなり、移動できずにその場にとどまるミクログリアが増加しました。さらにE15におけるミクログリアの分布を調べた結果、皮質板に存在するミクログリアが増加していました。以上のことから、髄膜や脳室下帯が産生するCXCL12をミクログリアがCXCR4を介して感知し、その産生源に向かって脳実質内を両方向に移動した結果、皮質板から不在となることが示唆されました(図3)。
では、ミクログリアが胎生中期に皮質板から一時的に抜け出すことにどのような意味があるのでしょうか。私たちは、ミクログリアの皮質板からの不在がニューロン成熟の適切な進行に必要なのではないか、という仮説を立てました。それを検証するために、ミクログリアを人工的に皮質板へと配置させ、ニューロンへの影響の有無を調べることにしました。様々な手法を用いてミクログリアを皮質板へと強制的に誘引したところ(皮質板内のニューロンが特異的にCXCL12を発現する遺伝子導入システムの利用や、脳から別途集めてきたミクログリアを皮質板に直接移植するなどの方法を用いて)、ミクログリア近傍のニューロンにおいて、将来の機能・性質を決める重要な分子群の発現パターンに乱れが生じていました。一般的に、ニューロンの運命は脳室帯や脳室下帯に存在する頃の神経前駆細胞の時点である程度決まる7ことが知られていますが、まだ完全には決まっておらず最終的な調節を皮質板に移動し終えた後で受けることが報告されています8。したがって、上述の結果は、過剰なミクログリアによってニューロンの成熟プロセスが乱れることを意味し、脳づくりの進行にはミクログリアが適切なタイミングで皮質板から退出する必要があることが初めて明らかとなりました。
さらに私たちは、ニューロンにおける遺伝子発現パターンの変化をきたすミクログリア由来分子の同定を進めました。培養下で用意した皮質板を構成するニューロンを、ミクログリア存在下、および、非存在下で培養した後にニューロンのみから全遺伝子を回収し、RNAシークエンス解析をおこないました。 両者の遺伝子発現を比較したところ、ミクログリアと共培養したニューロンにおいて、サイトカインの1型インターフェロン(IFN-I)とインターロイキン6(IL-6)によって活性化される細胞内シグナル経路に関連する分子群の発現が高まっていることがわかりました。 この結果に基づき、IFN-IとIL-6がニューロンの運命決定に関わる重要な分子群の発現変化に直接関わっている可能性を調べるため、細胞レベルおよび生体レベルで解析を行いました。これらの分子のはたらきを中和抗体により阻害したところ、ミクログリアによるニューロンの性質変化が抑えられたことから、ミクログリアから産生されるIFN-IとIL-6がニューロンの運命決定に関わる分子群の発現変化を促す要因であることが明らかとなりました(図4)。
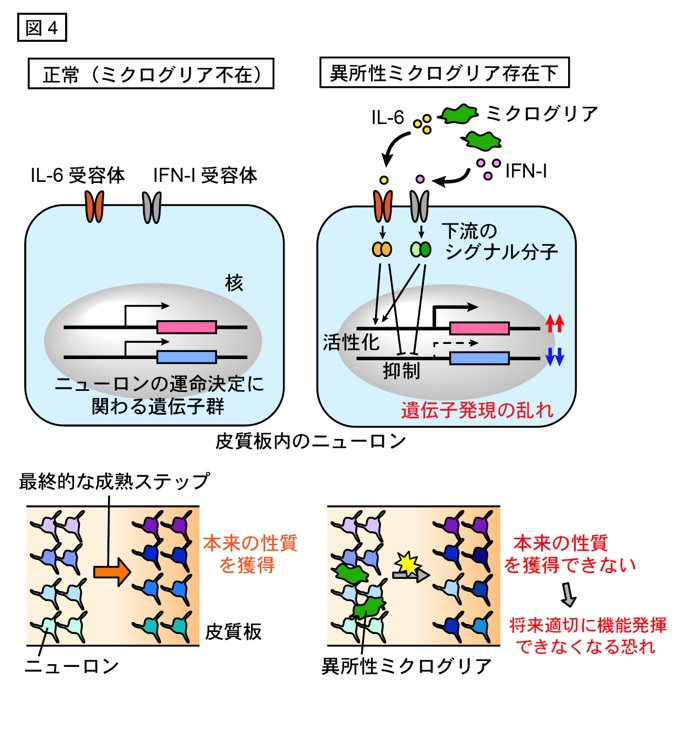
以上の結果から、ミクログリアは胎生中期にCXCL12に誘引され皮質板を一時的に抜け出し、ミクログリア由来の IFN-I や IL-6 によってニューロンの成熟ステップが乱されないよう巧みに分布を調節していることが示唆されました9。一つ一つのニューロンが生後の脳で正しく機能を発揮できるように、ミクログリアはこの時期にあえて皮質板から離れ、ニューロンの成熟を遠くから見守っていると考えられます。
今後の展開
本研究により、ミクログリアの正常脳における分布や移動のメカニズム、そして脳発生過程におけるその意義について詳細が明らかになりました。本成果は、ミクログリアとニューロン、アストロサイト、オリゴデンドロサイト、および、それ以外の細胞(血管等を構成する非神経系細胞)との細胞間ネットワークの理解や、胎生期にとどまらず生後から成体にわたる脳発達や機能成熟の理解にも大きく貢献すると考えます。一方で、感染症、低栄養・肥満等による母体の免疫活性化が胎児の統合失調症、自閉症等の発症リスクを高めることが近年報告されています。ミクログリアが母体炎症を感知することも示唆されていますが、ミクログリアがいかに性質を変え、脳を築く他の細胞との関係性にどのような影響が生じるのかについてはまだ明らかにされていません。本研究によって得られた「正常な脳におけるミクログリアの動態」に関する情報は、母体炎症時の細胞の動きの変化を見出すのに役立ち、胎児の脳発生異常を未然に防ぐための予防法や治療法の開発に繋がると期待されます。
参考文献
1) Matcovitch-Natan O et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science 353, aad8670 (2016).
2) Wake H et al. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J. Neurosci. 29, 3974–3980 (2009).
3) Paolicelli RC et al. Synaptic pruning by microglia is necessary for normal brain development. Science 333, 6048:1456–1458 (2011).
4) Hattori Y and Miyata T. Microglia extensively survey the developing cortex via the CXCL12/CXCR4 system to help neural progenitors to acquire differentiated properties. Genes Cells 23, 915–922 (2018).
5) Cunningham CL et al. Microglia regulate the number of neural precursor cells in the developing cerebral cortex. J. Neurosci. 33, 4216–4233 (2013).
6) Squarzoni P et al. Microglia modulate wiring of the embryonic forebrain. Cell Rep. 8, 1271–1279 (2014).
7) Molyneaux BJ et al. Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neurosci. 8, 427–437 (2007).
8) Kwan KY et al. SOX5 postmitotically regulates migration, postmigratory differentiation, and projections of subplate and deep-layer neocortical neurons. Proc. Natl. Acad. Sci. USA 105, 16021–16026 (2008).
9) Hattori Y et al. Transient microglial absence assists postmigratory cortical neurons in proper differentiation. Nat. Commun. 11, 1631 (2020).
「神経化学トピックス」では、神経化学のトピックを一般の方にもわかりやすくご紹介します。
※なお、目次記載の所属は執筆当時の所属となっております。