41. 神経障害性疼痛を緩和するミクログリアサブタイプ
はじめに
痛覚は、有害な刺激から身を守るために必要な生体警告信号です。しかし、がんや糖尿病、帯状疱疹後では、通常よりも痛みが強くなったり、触刺激でも痛みが出現するなど、刺激の強度や質に見合わない痛みが生じてしまいます。これらの痛みは、体性感覚神経系の損傷や病変などで発症するため神経障害性疼痛と呼ばれます。既存の鎮痛薬が効かないことも多く、そのメカニズムの解明と新しい治療法の開発が求められています。
末梢組織への刺激に相応しない痛みの原因として中枢神経系でのネットワーク異常が考えられています。私たちはこれまでの研究から、神経損傷に伴い脊髄後角で活性化したミクログリア細胞が神経障害性疼痛の原因の一つであることを示してきました1。その後、神経-グリア相互作用に注目した疼痛研究が数多くなされ、神経障害性疼痛の発症メカニズムの理解が深まってきました2。さらに近年、シングルセル遺伝子発現解析技術の発展に伴い、ミクログリアサブタイプの存在が明らかにされ、それら独自の機能や役割に関心が集まっています。しかし、神経障害性疼痛におけるミクログリアの多様性やサブタイプ独自の役割は全く分かっていませんでした。
神経障害性疼痛の発症‘後’に増加するCD11c陽性ミクログリア
本研究では、中枢神経疾患時に細胞数が増加するミクログリアとして報告されていたCD11c陽性サブタイプに注目しました3。CD11c陽性細胞でVenusを発現するマウスで神経障害性疼痛モデルを作製し、同マウスの脊髄におけるCD11c陽性細胞を組織学的に調べた結果、損傷した末梢神経が投射する脊髄領域においてその細胞数が増加することが分かりました。従来の研究対象であったミクログリア(CD11c陰性)の細胞数は損傷後7日目にピークを迎えますが、興味深いことに、CD11c陽性ミクログリア細胞数のピークは損傷後14日目となりました。CD11cを強く発現するCD11c強陽性ミクログリアは35日目にピークとなりました。その出現タイムコースが従来研究されてきたミクログリア(発症期に増加)とは明らかに異なることから、CD11c陽性ミクログリアはこれまで知られていなかった役割を有する可能性が考えられました(図1)。
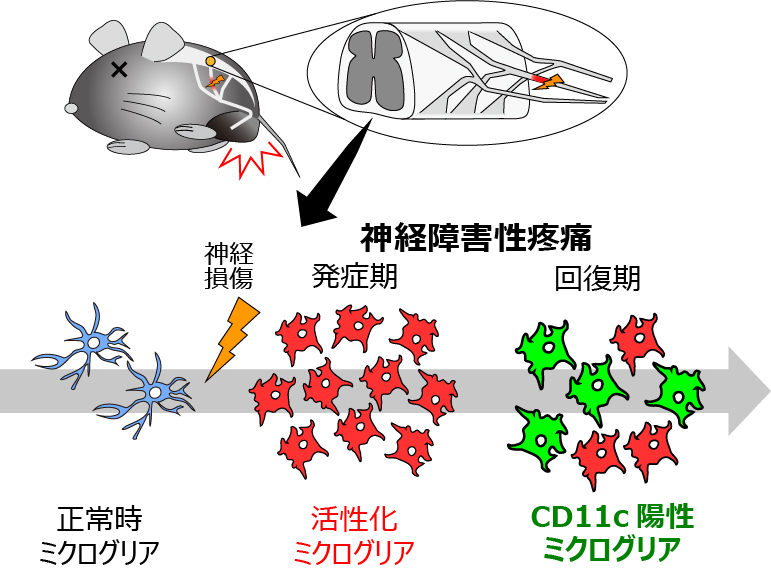
図1 CD11c陽性ミクログリアは疼痛の発症後に増加する
神経障害性疼痛モデルは、末梢神経の一部を切断することで作製した。CD11c陽性ミクログリアは、これまで考えられてきた時期より後期(疼痛の回復期)に活性化した。
CD11c陽性ミクログリアは神経障害性疼痛の自然回復に必要
マウスでの神経障害性疼痛は、マウスの足底皮膚にvon Freyフィラメントを押し当て、その刺激に対する逃避行動の過敏化を指標に評価します。この痛覚過敏に対するCD11c陽性ミクログリアの関与を調べるため、CD11c陽性ミクログリアを除去する方法を利用しました。具体的には、CD11c陽性ミクログリアにジフテリア毒素受容体を発現する遺伝子改変マウスの脊髄くも膜下腔内へジフテリア毒素を投与します。ジフテリア毒素は通常のマウス細胞には作用しないため、CD11c陽性ミクログリア選択的に細胞死を誘導できます。神経損傷後7日目や14日目にジフテリア毒素を投与しCD11c陽性ミクログリアを除去したところ、数日経過しても痛覚過敏に影響はありませんでした。したがって、CD11c陽性ミクログリアは疼痛の維持には関わらないことが分かりました。しかし、14日目以降、CD11c陽性ミクログリアを持続的に除去したところ、通常認められる疼痛の自然緩解が顕著に阻害されることを見いだしました(図2)。これらの結果は、CD11c陽性ミクログリアが疼痛状態からの回復に必要であることを示しています。そこで次に、この細胞がどのように増加し、またどのように痛みを緩和しているのかを調べることとしました。
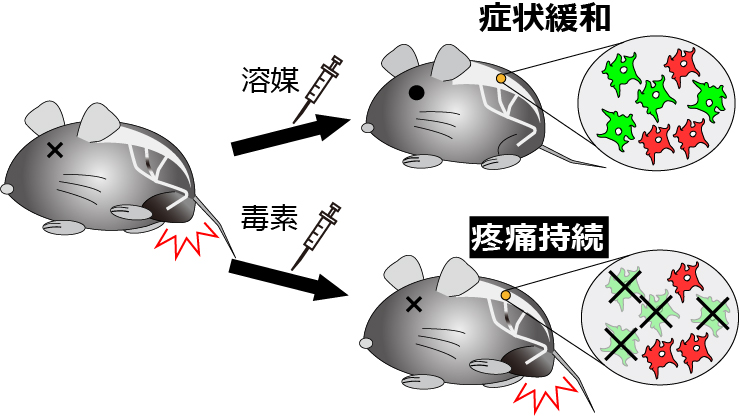
図2 CD11c陽性ミクログリアは神経障害性疼痛を緩和する作用を有する
遺伝子改変マウスにジフテリア毒素を投与することで、CD11c陽性ミクログリアを選択的に除去できる。溶媒を投与したマウスでは痛覚過敏が自然緩解するが、CD11c陽性ミクログリアを持続的に除去したマウスでは疼痛症状が持続した。
ミエリンの貪食によってCD11c陽性ミクログリアが出現する
神経障害性疼痛モデルマウスの脊髄後角からCD11c陽性ミクログリアと陰性ミクログリアをFACSにて分取し、RNAシーケンス法で両者の遺伝子発現を網羅的に解析した結果、数多くの遺伝子発現に差異が認められました。CD11c陽性ミクログリアでは、細胞貪食に関わる遺伝子の発現が高く、実際に同ミクログリアはミエリンデブリの貪食能が高いことが分かりました。加えて、精製したミエリンそのものを脊髄後角実質内に注入すると、ミクログリアがそれを貪食し、CD11c陽性となりました。そこで、CD11c陽性ミクログリアで発現が高かったAXLに注目しました。AXLは貪食関連分子であるTAMファミリー分子の一つです。AXL遺伝子欠損マウスではCD11c陽性ミクログリアが少なく、疼痛症状も遷延化しました。以上の結果から、CD11c陽性フェノタイプに移行するトリガーのひとつがミエリンデブリの貪食であることが示唆されました(図3)。
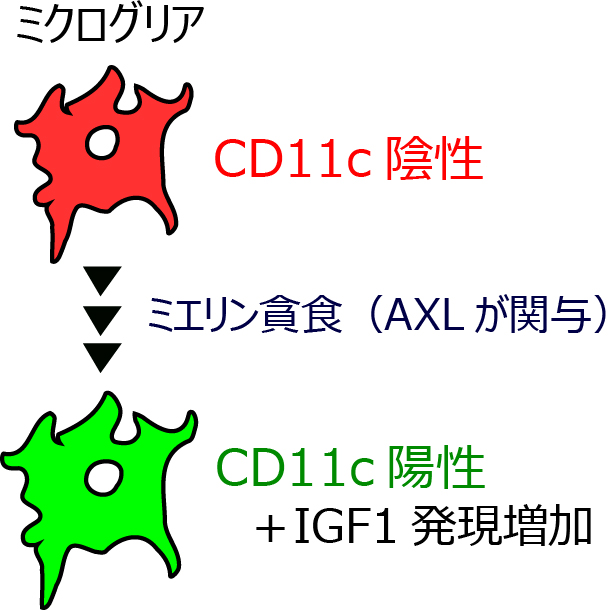
図3 CD11c陽性ミクログリアは貪食を介して出現し、IGF1を介して疼痛を緩和する
CD11c陽性ミクログリアの出現にはミエリンの貪食やAXLが関与し、IGF1の発現も増加する。IGF1は疼痛の緩和作用を有していた。
CD11c陽性ミクログリア由来のIGF1が痛みの緩和に寄与する
CD11c陽性ミクログリアで高発現していた液性因子として、インスリン様成長因子1(insulin-like growth factor-1; IGF1)がありました。IGF1の発現は、疼痛の自然回復のタイムコースと相関し、CD11c陽性ミクログリアの機能発現に重要な因子である可能性が考えられました。そこで、IGF1遺伝子を欠損する遺伝子改変マウスを複数系統(CD11c陽性細胞もしくはミクログリア細胞選択的に欠損)用いて、それらの神経障害性疼痛モデルを作製しました。その結果、痛覚過敏の発症には影響がなかったものの、自然回復が見られないことが分かりました。以上の結果からCD11c陽性ミクログリアにおいては、IGF1が発現増加し、それがアロディニア症状の緩和に重要な役割を果たしていることが示されました(図3)。
おわりに
これまでの研究から、脊髄のミクログリアは神経損傷後早期に活性化し、神経障害性疼痛の発症に重要であると考えられてきました。今回の研究では、疼痛の発症後に増加するCD11c陽性ミクログリアを見いだし、さらにこのサブタイプが疼痛を緩和する役割を持つことを明らかにしました4。本研究の成果から、CD11c陽性ミクログリアを増やすという新しい慢性疼痛治療戦略の可能性を提示できました。今後は、CD11c陽性ミクログリア増加メカニズムの解明やその増生法の確立、IGF1の疼痛抑制メカニズムに関する研究に取り組む必要があると考えています。
参考文献
- Tsuda M, Shigemoto-Mogami Y, Koizumi S, Mizokoshi A, Kohsaka S, Salter MW, Inoue K. P2X4 receptors induced in spinal microglia gate tactile allodynia after nerve injury. Nature, 424, 778–783 (2003). doi:10.1038/nature01786
- Inoue K, Tsuda M. Microglia in neuropathic pain: cellular and molecular mechanisms and therapeutic potential. Nat Rev Neurosci, 19, 138–152 (2018). doi:10.1038/nrn.2018.2
- Benmamar-Badel A, Owens T, Wlodarczyk A. Protective microglial subset in development, aging, and disease: lessons from transcriptomic studies. Front Immunol, 11, 430 (2020). doi:10.3389/fimmu.2020.00430
- Kohno K, Shirasaka R, Yoshihara K, Mikuriya S, Tanaka K, Takanami K, Inoue K, Sakamoto H, Ohkawa Y, Masuda T, Tsuda M. A spinal microglia population involved in remitting and relapsing neuropathic pain. Science, 376, 86–90 (2022). doi:10.1126/science.abf6805
このページの目次
「神経化学トピックス」では、神経化学のトピックを一般の方にもわかりやすくご紹介します。
※なお、目次記載の所属は執筆当時の所属となっております。