24. マウス発生期大脳皮質において、リーリンはN-カドヘリン依存的な神経細胞接着を一過的に促進する
はじめに
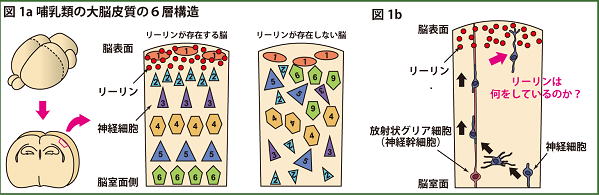
私たちの脳の知覚、思考、記憶などの高次機能を司る大脳皮質(大脳新皮質)では、神経細胞がきれいな6層構造を作っています(図1a)。この6層の細胞層を構成する興奮性の神経細胞は、脳の深部にある脳室の近くで誕生します。生まれた神経細胞は、放射状グリア細胞と呼ばれる神経幹細胞が脳室面から脳表面へと伸ばす長い突起“放射状グリア線維”を足場として利用し、脳表面の辺縁帯直下まで移動してその移動を終えます(図1b)。
リーリンは辺縁帯に存在するCajal Retzius細胞から細胞外に分泌される糖タンパク質であり、リーリンが欠損するリーラーマウスでは、上記の層構造がうまく作られず全体として逆転するという大きな表現型を示すことが知られています(図1a)。リーリンは、これまで神経細胞の移動を終わらせるための単純な移動停止シグナルであると考えられてきました。
しかしながら我々の研究グループは、リーリンを発生期脳内で移動途中に強制的に発現させると、その周囲に移動中の神経細胞が集まって凝集塊を作ることを発見し、以前報告しました(文献1)(以下では「リーリンが誘導するin vivoの細胞凝集塊」と呼びます)。ところがこの以前の研究では、リーリンが直接神経細胞間の接着力を強めて凝集させているのか、あるいは、リーリンが神経細胞の周囲の環境に働きかけ、神経細胞が周囲から反発されるなどして結果として凝集しているのかは区別がついていませんでした。
リーリンは神経細胞の凝集を直接誘導する
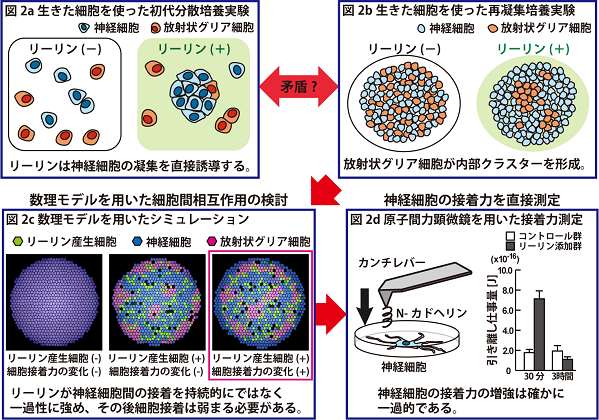
そこでまず、リーリンが神経細胞の凝集を直接誘導するかを調べる為に、神経細胞を分散して初代培養を行い、培地にリーリンを添加しました(文献2)。その結果、リーリンは分散した神経細胞に直接作用して凝集させる機能を持っていることがわかりました(以下では「リーリンによるin vitroの細胞凝集」と呼びます)(図2a)。神経細胞には、リーリンを受け取るリーリン受容体と、その下流でリーリンシグナルを細胞内に伝達するDab1という分子が発現していますが、Dab1を欠損したマウスで同様の実験を行っても細胞凝集は見られませんでした。この結果から、リーリンによる神経細胞の凝集はDab1経路を介していることもわかりました。
リーリンは細胞接着分子を制御することが既に知られていますが、辺縁帯にはN-カドヘリンが強く検出されること、また、リーリンが誘導するin vivoの細胞凝集塊の中心部でもN-カドヘリンが検出されることから、リーリンが神経細胞の凝集を誘導する際に制御する接着分子の候補としてN-カドヘリンに着目しました。N-カドヘリンのノックダウンやドミナントネガティブ体の強制発現による機能阻害を行ったところ、リーリンによるin vivoおよびin vitroの細胞凝集が阻害されたことから、リーリンが誘導する細胞凝集にはN-カドヘリンが必要であることが示されました(文献2)。
リーリンは神経細胞の接着を一過的に強める
一方、発生期大脳皮質の細胞を分散後、試験管内で回旋させながら再凝集させる培養系において、リーリン存在下では神経細胞と放射状グリア細胞とが再凝集塊内で選別配置されることを見いだしました(文献2)。上記のリーリンによる神経細胞凝集実験の結果からは、神経細胞が再凝集塊内の中心付近で集まってクラスターを作ることが予想されましたが、意外なことに結果は逆でした。すなわち、再凝集塊の中心近くでは神経細胞ではなく、むしろ放射状グリア細胞が集まってクラスターを形成して神経細胞と選別されていました(図2b)。そこで、いかなる細胞間相互作用を想定すれば、このような選別パターンが実現できるのかを予測する目的で、数理モデルによる検討を行いました。数理モデルでシミュレーションを行い検討した結果、再凝集培養実験の選別パターンを実現するためには、リーリンが神経細胞間の接着を持続的にではなく一過性に強め、その後細胞接着が弱まる必要があるということが示唆されました(図2c)。
そこで次に、原子間力顕微鏡を用いてN-カドヘリンタンパク質に対する神経細胞の接着力を直接測定しました(文献2)。神経細胞をリーリンで刺激したところ、刺激後30分ではN-カドヘリンタンパク質との接着が強まる一方で、刺激後3時間では接着力が元に戻る結果が得られました(図2d)。
神経細胞の一過的な接着力の増強が、神経細胞の最終配置に重要である。
そこでさらに、神経細胞間の細胞接着が増強されたまま減弱しない場合の大脳皮質の層形成への影響を調べました(文献2)。すなわち、胎生14.5日齢のマウス胎児に子宮内電気穿孔法を用いて遺伝子導入を行い、細胞接着に直接関与すると予測される細胞膜上のN-カドヘリン分子の安定化を行って、N-カドヘリンを介した接着が強いまま弱まらないようにしてみました。その結果、生後7.5日目で神経細胞の配置の乱れが観察されました。以上の結果より、リーリンによって増強された神経細胞間のN-カドヘリン依存的な接着は、その後弱くなることが重要であることが示唆されました。
おわりに
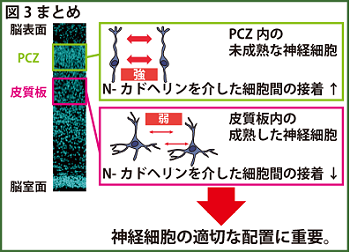
今回見出された現象は、リーリンが本来産生される辺縁帯の直下において、移動を終えたばかりの未熟な神経細胞が帯状に密に並ぶprimitive cortical zone (PCZ)と呼ばれる領域(文献3)で、神経細胞が一過的に高密度で局在するメカニズムと関係があると予測されます。未熟な神経細胞は24時間程度でPCZを離れ成熟した神経細胞になりますが、このタイミングで神経細胞間の接着が弱まる可能性が示唆されます(図3)。
図の説明
図1a, b 哺乳類の脳の神経細胞が作る 6 層構造。この層構造が正しく作られるために、リーリンと呼ばれるタンパク質が必須であることが知られています。
図2a 分散して培養した大脳皮質細胞の培地にリーリンを添加すると、神経細胞の凝集が誘導されました。
図2b 胎生期大脳皮質の細胞を分散させ、試験管内で旋回させながら再凝集させる培養系にリーリンを添加したところ、神経細胞よりもむしろ放射状グリア細胞がクラスターを形成しているように観察されました。
図2c シミュレーションを行って図2bの選別パターンを再現できる条件を検討した結果、リーリンは神経細胞間の接着を持続的にではなく一過的に増強する可能性が示唆されました。
図2d 原子間力顕微鏡を用いて細胞接着力を測定したところ、リーリンによる神経細胞の接着力の増強は確かに一過的であることがわかりました。
図3 神経細胞が移動を終えて層構造を形成し始める部位であるPCZでは、リーリンによって細胞間の接着が一過的に増強し、その後神経細胞が成熟すると接着は弱くなることが示唆されました。この細胞間の接着が強くなったまま弱くならないと、層構造は異常になってしまうこともわかりました。
参考文献
1) Kubo, K., Honda, T., Tomita, K., Sekine, K., Ishii, K., Uto, A., Kobayashi, K., Tabata, H., Nakajima, K. Ectopic Reelin induces neuronal aggregation with a normal birthdate-dependent "inside-out" alignment in the developing neocortex. J. Neurosci. 30, 10953-10966. (2010).
2) Matsunaga, Y., Noda, M., Murakawa, H., Hayashi, K., Nagasaka, A., Inoue, S., Miyata, T., Miura, T., Kubo, K.I., Nakajima, K. Reelin transiently promotes N-cadherin-dependent neuronal adhesion during mouse cortical development. Proc. Natl. Acad. Sci. USA 114, 2048-2053. (2017).
3) Sekine, K., Honda, T., Kawauchi, T., Kubo, K., Nakajima, K. The outermost region of the developing cortical plate is crucial for both the switch of the radial migration mode and the Dab1-dependent "inside-out" lamination in the neocortex. J. Neurosci. 31, 9426-9439. (2011).
「神経化学トピックス」では、神経化学のトピックを一般の方にもわかりやすくご紹介します。
※なお、目次記載の所属は執筆当時の所属となっております。