42. ミクログリアはホスファチジルセリン依存的に成体新生ニューロンのシナプスを貪食する
はじめに
哺乳類の脳で産生された新しいニューロンは、嗅球や海馬で他のニューロンとシナプスを形成することで高度な神経回路を形成します。この成体新生ニューロンの成熟過程には、中枢神経系の貪食細胞であるミクログリアが重要な役割を果たします。ミクログリアは死細胞や発達期のシナプスを貪食する1)~3)ことが分かっていますが、成体新生ニューロンの成熟過程におけるシナプス貪食についてはあまり報告されておらず、分子メカニズムも不明でした。私たちは、このメカニズムを明らかにするため、ホスファチジルセリン(PS)という細胞膜を構成するリン脂質に着目し、ミクログリアが成体新生ニューロンのシナプスを貪食する仕組みを調べました。
成体新生ニューロンのシナプス貪食とPSの露出
まず、電子顕微鏡4) 5)でミクログリアを詳しく観察したところ、ミクログリアが成体新生ニューロンのシナプスを貪食している様子が観察されました。
PSは、通常は細胞膜の内側に存在しますが、死細胞では細胞膜の外側に露出し、貪食を促進することが分かっています。そこで、細胞膜の外側のPSを検出する試薬を用いて、成体マウスの脳内でのPSの分布を調べました。その結果、死細胞だけでなく、シナプスにおいても、PSが細胞膜の外側に露出していることが観察されました。
シナプス貪食過程におけるPSの機能
次に、ミクログリアによる成体新生ニューロンのシナプス貪食におけるPSの機能を調べるために、新しい遺伝子改変マウス(D89Eマウス)を作製しました。このマウスでは、PSに結合するオプソニンであるMFG-E8の変異体(MFG-E8D89E)を用いることで、細胞膜の外側に露出したPSをマスクし、PS依存的な貪食を阻害することができます。ミクログリアと新生ニューロンのシナプスの様子を観察し、D89Eマウスと野生型マウスとで比較したところ、D89Eマウスでは、野生型マウスに比べて、シナプス上でのミクログリアの膜の広がりや貪食が阻害されました。このことから、PSはミクログリアの膜を広げることで、シナプス貪食を促進することが示唆されました(図1)。
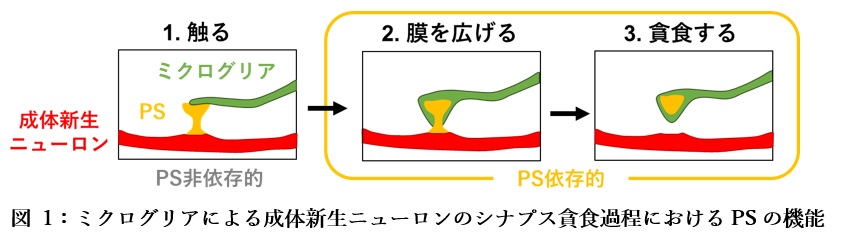
PS依存的なシナプス貪食の意義
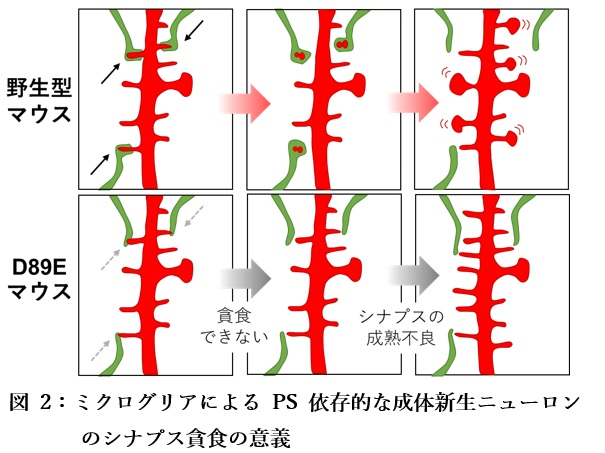
最後に、D89EマウスでPS依存的なシナプス貪食を阻害すると、シナプスの密度や機能に影響が出るかを調べました。海馬の成体新生ニューロンのシナプス密度をD89Eマウスと野生型マウスで比較したところ、D89Eマウスでは未熟なシナプスの密度は増加したものの成熟したシナプスの密度は減少しました(図2)。さらに、シナプスの機能を示す電気生理学的な特徴にも異常が見られることが分かりました。このことから、ミクログリアによるPS依存的な成体新生ニューロンのシナプス貪食は、海馬新生ニューロンのシナプス成熟に関与していることが示唆されました。
まとめと展望
私たちは、PSをマスクする新しい遺伝子改変マウスを用いることにより、ミクログリアがPS依存的に成体新生ニューロンのシナプスを貪食することを明らかにしました6)。最近の研究で、人でも新生児期には、新しくニューロンが産生されることがわかっており、生後の脳発達に重要だと考えられます。また、自閉症モデルマウスや自閉症患者の死後脳で、ミクログリアの形態異常やシナプス密度の増加が報告されていることから、ミクログロリアによるシナプス貪食の異常が自閉症の病態と関連している可能性が考えられます。今後、PSとミクログリアによるシナプス貪食の関連をさらに詳しく調べることで、新たな治療法の開発につながる可能性が考えられます。
参考文献
- Scott-Hewitt N, Perrucci F, Morini R, Erreni M, Mahoney M, Witkowska A, Carey A, Faggiani E, Schuetz LT, Mason S, Tamborini M, Bizzotto M, Passoni L, Filipello F, Jahn R, Stevens B, and Matteoli M. Local externalization of phosphatidylserine mediates developmental synaptic pruning by microglia. EMBO J. 39, e105380 (2020) DOI: 10.15252/embj.2020105380.
- Li T, Chiou B, Gilman CK, Luo R, Koshi T, Yu D, Oak HC, Giera S, Johnson-Venkatesh E, Muthukumar AK, Stevens B, Umemori H, and Piao X. A splicing isoform of GPR56 mediates microglial synaptic refinement via phosphatidylserine binding. EMBO J. 39, e104136 (2020) DOI: 10.15252/embj.2019104136.
- Park J, Choi Y, Jung E, Lee SH, Sohn JW, and Chung WS. Microglial MERTK eliminates phosphatidylserine-displaying inhibitory post-synapses. EMBO J. 40, e107121 (2021) DOI: 10.15252/embj.2020107121.
- Matsumoto M, Sawada M, GarcÍa-González D, Herranz-Pérez V, Ogino T, Nguyen HB, Thai TQ, Narita K, Kumamoto N, Ugawa S, Saito Y, Takeda S, Kaneko N, Khodosevich K, Monyer H, García-Verdugo JM, Ohno N, and Sawamoto K. Dynamic changes in ultrastructure of the primary cilium in migrating neuroblasts in the postnatal brain. J. Neurosci. 39, 9967 (2019) DOI: 10.1523/JNEUROSCI.1503-19.2019.
- Sawada M, Ohno N, Kawaguchi M, Huang S, Hikita T, Sakurai Y, Nguyen HB, Thai TQ, Ishido Y, Yoshida Y, Nakagawa H, Uemura A, and Sawamoto K. PlexinD1 signaling controls morphological changes and migration termination in newborn neurons. EMBO J. 37, e97404 (2018) DOI: 10.15252/embj.201797404.
- Kurematsu C, Sawada M, Ohmuraya M, Tanaka M, Kuboyama K, Ogino T, Matsumoto M, Oishi H, Inada H, Ishido Y, Sakakibara Y, Nguyen HB, Thai TQ, Kohsaka S, Ohno N, Yamada MK, Asai M, Sokabe M, Nabekura J, Asano K, Tanaka M, and Sawamoto K. Synaptic pruning of murine adult-born neurons by microglia depends on phosphatidylserine. J. Exp. Med. 219, e20202304 (2022) DOI: 10.1084/jem.20202304.
「神経化学トピックス」では、神経化学のトピックを一般の方にもわかりやすくご紹介します。
※なお、目次記載の所属は執筆当時の所属となっております。