27. 超解像度解析による成長円錐の新たな膜小胞動態の発見
脳の働きはシナプス(神経細胞同士の機能的結合)での、信号の送信で成り立っていますが、シナプスは胎生期に遺伝子の働きによって自動的に作られます。この時、神経細胞から軸索と言う突起が伸び、その先端に運動性の高い構造体が作られて標的となる神経細胞まで伸びていきます。この特殊構造を成長円錐(growth cone)と呼び、成長円錐は機関車のように目的地まで常に決まった経路で神経突起を牽引し、標的の細胞を認識すると停止してシナプスを形成します。従って、成長円錐は神経回路形成の中核となる、決定的に重要な意義を持ちます(図1)。
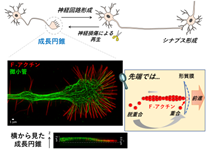
図1 成長円錐と神経回路形成. 成長円錐が細胞骨格などの働きで標的まで伸びる
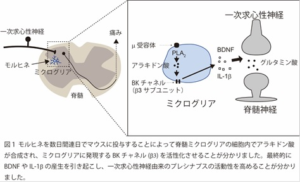
上述の働きを成長円錐が持つため、その情報はこの構造の中の分子群が担っていると考えられます。成長円錐を構成する成分は、アクチン繊維や微小管を含む「細胞骨格」、神経細胞の表面の膜と膜小胞の「膜成分」の2つに大別されます。これらの成分に蛍光タンパク質と結合させれば原理的にライブイメージングが出来、分子群の動態が解明できるはずですが、実際には成長円錐内部は非常に混み合っており、原理的に分解能200 nmが限界(Abbeの限界)である通常の光学顕微鏡では明確に見られません。
最近、光学顕微鏡でありながらAbbe限界の壁を破る方法が開発され、100~20nmの分解能をもつ「超解像(度)顕微鏡」ができました(2014年ノーベル化学賞)。例えば「細胞骨格」と「膜小胞」の双方に着目し、同時に複数の分子を可視化するには光学顕微鏡を使うしかなく、われわれは超解像(度)顕微鏡の一種で、構造化照明顕微鏡(SIM)を導入して解析を行いました(図2)。
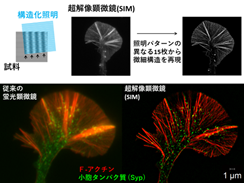
図2 SIMの原理と成長円錐の分子観察. パターンの重ね合せから鮮明な像が得られる
SIMは縞模様の照明を用いることで、試料の微細構造の情報を含む蛍光像に干渉縞が生じ、計算によって画像に再現できます。特に動画を得ることが出来るのは他の超解像度顕微鏡に無い特長で、超高精細の画像を元に分子の動態が解明できます。
SIM解析の結果、アクチン細胞骨格が伸長してアクチンの束形成時に、近傍で細胞膜の一部が細胞内に取り込まれて輸送小胞が生まれる様子を、動画で初めて捉えることに成功しました(図3)。この取込みはエンドフィリンと呼ばれる、膜の曲率を作り出すタンパク質が集合することが証明されました。アクチンの束形成を阻害すると、成長円錐のアクチン束が減るだけに留まらず、先端にエンドフィリンが集合せず、神経先端ではアクチン細胞骨格の伸長によって局所的な細胞膜取込みが誘導されていると結論できます。阻害実験を行って神経成長にエンドフィリンが必要であることも証明でき、超解像度顕微鏡による成長円錐構造の3次元再構成の結果、スタンダードな細胞膜取込みの誘因因子として広く知られているクラスリンは成長円錐の底面に多いのに対し、エンドフィリンはZ軸側(上側)の成長円錐表面の膜に多く集積することが初めて明らかにできました(図3)。
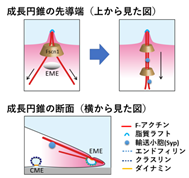
図3 成長円錐の超解像度観察の結果. 空間的に全く異なる機構の小胞動態が証明された
これらの結果から、神経が伸びながら先端表面の細胞膜を積極的に取り込むことによって、細胞外環境を探る成長円錐のメカニカルな仕組みが解明できました(文献1)。成長円錐が動く仕組みを明らかにするため、当研究グループはプロテオーム解析で成長円錐を構成する種々のタンパク質同定に成功したものの(文献2)、SIM導入以前は、上述の理由で分子同士の相互関係がはっきりせず、機能的意義がよくわかりませんでした。今回の研究で機能未知の分子でも神経成長における意義がより明確に理解できました(例えばシグナル伝達の場として知られる、膜ドメインの脂質ラフトと機能未知の蛋白質GPM6aの関係性など)(文献1、3)。
成長円錐の膜上にはガイダンス分子を受け取るための様々な受容体が存在し、成長円錐内ではそれらの働きに対応して、細胞骨格を再編して形態を変化させ、進行方向、前進または停止を制御しています。今回のわれわれの発見をさらに発展させ、神経成長の分子機構を明らかにすることで、神経疾患への新たな治療戦略への寄与が可能となるでしょう(文献4)。
文献
1)Nozumi M, Nakatsu F, Katoh K, Igarashi M (2017) Coordinated movement ofvesicles and actin bundles during nerve growth revealed by superresolutionmicroscopy. Cell Rep 18: 2203-2216.
2)Nozumi M, Togano T, Takahashi-Niki K, Lu J, Honda A, Taoka M, Shinkawa T, KogaH, Takeuchi K, Isobe T, Igarashi M (2009) Identification of functional markerproteins in the mammalian growth cone. Proc Natl Acad Sci USA 106: 17211-17216.
3)Honda A, Ito Y, Takahashi-Niki K, Matsushita N, Nozumi M, Tabata H, Takeuchi K,Igarashi M (2017) Extracellular signals induce glycoprotein M6a clustering oflipid rafts and associated signaling molecules. J Neurosci 37: 4046-4064.
4)Nozumi M, Igarashi M (2017) Vesicular movements of the growth cone. NeurochemInt (Sep 26. pii: S0197-0186(17)30368-6. doi: 10.1016/j.neuint.2017.09.011).
「神経化学トピックス」では、神経化学のトピックを一般の方にもわかりやすくご紹介します。
※なお、目次記載の所属は執筆当時の所属となっております。